
Biodiversité génétique
- Fiche de cours
- Quiz et exercices
- Vidéos et podcasts
Au cours de la méiose (formation des gamètes mâles et femelles), se produisent des brassages génétiques qui conduisent à une augmentation de la diversité génétique. Ils se combinent avec le brassage génétique de la fécondation.
Toutefois, le génome peut subir d’autres modifications génétiques qui vont conduire à l’émergence d’espèces présentant des caractères nouveaux. Nous allons étudier ces modifications.
C’est le cas de H. annuus (tournesol annuel), H. petiolaris (tournesol à long pétiole) et H. anomalus (tournesol anormal) qui vivent à l’ouest du pays.
Ces trois espèces possèdent le même nombre de chromosomes (2n = 34).
L’étude chromosomique de ces trois espèces a montré des similitudes :
- les chromosomes 1 à 6 sont identiques entre les trois espèces ;
- les chromosomes 10, 11, 15 et 16 sont identiques entre l’espèce H. petiolaris et H. anormalus alors que le chromosome 16 est le même entre l’espèce H. annuus et H. anormalus.
L’espèce H. anormalus possède aussi dans son génome des chromosomes présentant leur propre remaniement n’ayant pu se réaliser qu’après au moins trois cassures, trois fusions et une duplication à partir des chromosomes des deux autres espèces.
Ces observations suggèrent que l’espèce H. anormalus a hérité son génome du remaniement chromosomique des deux autres espèces. C’est donc une espèce hybride.
• Comment cette hybridation a-t-elle pu se réaliser ?
Les deux espèces parents sont des espèces sympatriques, c’est-à-dire des espèces coexistant dans un même territoire.
Leur floraison est légèrement décalée et les pollinisateurs sont communs aux deux espèces. Dans ces conditions, l’hybridation peut avoir lieu. Elle donne tout d’abord naissance à des hybrides de première génération F1 semi-stériles. Puis, au fil des générations, seules subsistent les hybrides présentant des recombinaisons chromosomiques favorables. Ils sont fertiles entre eux. On aboutit ainsi à leur isolement reproducteur et donc à l’émergence d’une nouvelle espèce présentant des caractéristiques différentes des espèces d'origine.
Dans le cas du tournesol anormal, il s’acclimate mieux aux milieux très secs.
Des simulations informatiques ont montré que la stabilisation de l’espèce hybride est rapide (25 à 60 générations seulement). La reconstitution en serre de cette hybridation conduit à la naissance d’espèces très proches génétiquement de H. anormalus.
On pense que ce processus peut survenir naturellement sans l’intervention de l’Homme.
• En quoi consiste la polyploïdisation ?
On sait qu’au cours de leur cycle de reproduction sexuée, les espèces passent d’un stade haploïde (n chromosomes) à un stade diploïde (2n chromosomes). Suivant l’espèce, la phase haploïde prédomine sur la phase diploïde ou l’inverse.
Dans le cas d’une espèce polyploïde, le nombre de chromosomes est le multiple d’un nombre supérieur à 2 (3n triploïdie ; 4n tétraploïdie, …).
Exemples de polyploïdie :
| Pomme de Terre | 4 × 12 = 48 chromosomes |
| Banane | 3 × 11 = 33 chromosomes |
| Cacahuète | 4 × 10 = 40 chromosomes |
| Tabac | 4 × 12 = 48 chromosomes |
| Coton | 4 × 13 = 52 chromosomes |
| Blé tendre | 6 × 7 = 42 chromosomes |
| Fraise | 8 × 7 = 56 chromosomes |
| Canne à sucre | 8 × 10 = 80 chromosomes |
• Comment se forme une espèce polyploïde ?
Au cours de la méiose, les chromosomes d’une même paire se séparent en deux lots identiques durant la première division méiotique. Si la membrane cellulaire ne se reforme pas entre les deux cellules, la cellule produite comporte 2n chromosomes et conduit à la production de gamètes à 2n chromosomes au lieu de n chromosomes.
Ce mécanisme peut être provoqué artificiellement par l’Homme chez les végétaux. Il utilise notamment la colchicine qui bloque la reconstruction de la membrane cellulaire. En effet, la polyploïdie est associée à des phénotypes de gigantisme cellulaire et morphologique qui permet d’optimiser la production végétale.
Il existe naturellement des espèces végétales :
- Autopolyploïdes qui se forment par duplication des chromosomes au sein d’une même espèce : la pomme de terre (4n = 48), la banane (3n = 33).
- Allopolyploïdes qui se forment après un croisement entre deux ou plusieurs espèces : le tabac (4n = 48), la fraise (8n = 56), le blé tendre (6n = 42).
Ce phénomène est beaucoup plus rare parmi les espèces animales. On cite cependant l’existence d’une espèce de Xénope (X. laevis) qui est une espèce allo-tétraploïde possédant 36 chromosomes. Si on la compare à l’espèce X. tropicalis diploïde on constate que la taille des individus mâles et femelles est beaucoup plus grande ainsi que la taille des œufs. De plus, ils ont une durée de vie plus longue.
On connaît aujourd’hui la séquence complète du génome humain. En plus des séquences codant pour des gènes connus et fonctionnels, on dénombre un grand nombre de séquences qui présentent des homologies avec des séquences virales (8 % seraient composées de vestiges rétroviraux).
Ainsi, la biodiversité du génome d’une espèce pourrait résulter de l’intégration stable de séquences virales. Au sein du génome de l’Homo sapiens, on peut citer l’exemple du gène codant la syncitine. C’est une glycoprotéine d’enveloppe rétrovirale (HERV-W) capable d’induire les fusions membranaires indispensables à l’entrée du virus dans la cellule cible.
Cette protéine est fortement produite par les cytotrophoblastes, cellules impliquées dans la formation du placenta. Elle permet la formation du syncytiotrophoblaste, siège des échanges materno-fœtaux.
C’est au 19e siècle que sont découverts les gènes homéotiques ; ils contrôlent la mise en place des organes et appendices. Leur mutation va perturber l’organisation spatiale de l’organisme et la morphologie des organes et appendices.
La drosophile est le premier modèle animal utilisé pour étudier la fonctionnalité de ces gènes. On identifie 8 gènes Hox répartis en 2 groupes (le complexe Antennapedia et le complexe Ultrabithorax) présents sur le chromosome 3. Ils déterminent la position des segments le long de l’axe antéro-postérieur de la drosophile. Au sein des complexes, les gènes sont disposés dans le même ordre de leur expression au sein de l’embryon.
La mutation du gène antennapedia conduit au positionnement de pattes à la place des antennes.
La comparaison des séquences de différents génomes a montré que les gènes Hox se retrouvent dans toutes les espèces animales testées. Les homologies de séquences sont importantes, jusqu’à 100 % parfois pour les homéodomaines de drosophile et de vertébrés.
Chez les vertébrés, les gènes Hox sont disposés en 4 complexes répartis sur 4 chromosomes différents. L’ordre des gènes au sein de ces complexes est aussi en relation avec leur ordre d’expression dans l’embryon. Par contre, l’identité de chaque segment est conférée par une combinaison d’expression de plusieurs gènes Hox qui lui est propre.
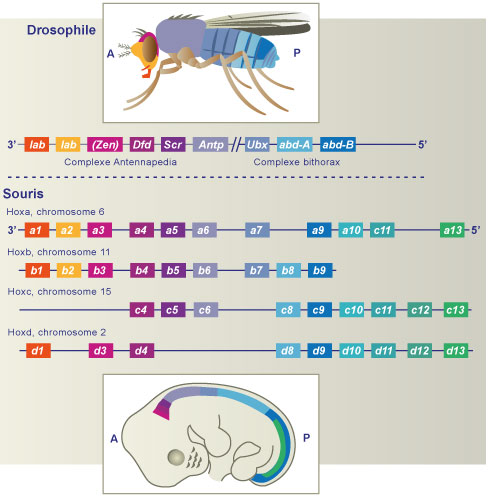
Les gènes homéotiques forment donc une famille multigénique issus de processus de duplication, crossing-over voire même polyploïdisation (duplication de tout le génome). Ils sont donc dits homologues et proviennent tous d’un même gène ancestral.
Le gène Pax6, commun à la drosophile et à l’Homme est impliqué dans la construction de l’œil. Sa mutation chez l’Homme conduit à une malformation de l’iris : l’aniridie. Chez la drosophile, elle conduit à une absence de l’œil. Lorsque l’on transfert le gène Pax6 humain à une cellule œuf de drosophile, on aboutit à la reconstruction de l’œil.
• Comment peut-on expliquer une diversité morphologique au sein des êtres vivants alors qu’elle dépend de gènes homologues ?
La fonction des gènes homéotiques dépend de l’endroit et de la période pendant laquelle ils sont exprimés au cours du développement embryonnaire.
Les gènes Hox6c et Hox8c sont exprimés dans les segments thoraciques :
- La limite antérieure de Hox6c se situe au niveau de la transition cervical / thoracique et détermine la position des pattes antérieures.
- La limite postérieure de Hox8c se situe au niveau de la patte postérieure.
Chez les serpents et notamment le python, la limite antérieure de la zone d’expression de Hox6c et Hox8c se situe dans la région crânienne. Ceci a pour conséquence l’apparition de côtes sur toutes les vertèbres et donc la disparition du cou.
Cette modification de l’expression spatiale des gènes Hox serait à l’origine de la disparition des membres antérieurs.
La disparition des membres postérieurs résulterait d’un mécanisme différent : absence d’un signal d’activation du développement du membre.
Le crâne du chimpanzé présente une face plus saillante « en museau » et un crâne proportionnellement moins globuleux ainsi qu’un trou occipital en position arrière.
L’Homme présente en fait les caractères juvéniles du crâne du chimpanzé : front bombé, arrière du crâne arrondi, face petite et placée sous le crâne, trou occipital en position centrale.
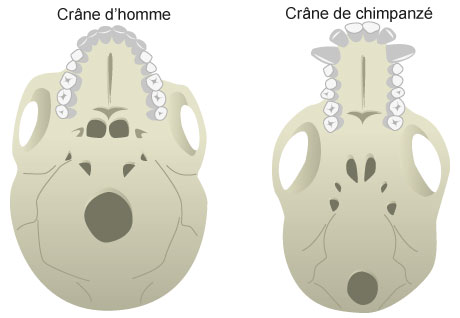
Tout se passe comme si la croissance du crâne humain s’était ralentie au cours du développement fœtal chez l’Homme. De plus, on constate que la phase embryonnaire au cours de laquelle les neurones se multiplient dure 2 semaines chez le chimpanzé contre 8 semaines chez l’Homme.
Le rallongement de cette phase du développement serait à l’origine du développement du cortex cérébral.
Ainsi, la modification de la chronologie de l’apparition des caractères fondamentaux au cours du développement embryonnaire peut aussi être à l’origine de la diversification du monde du vivant. Cette chronologie apparaît être sous le contrôle de gènes homéotiques.
Les allèles apparaissent au cours de l’évolution par mutation. Les nouveaux gènes proviennent de duplication de gènes ancestraux dont la séquence évolue.
La théorie de la sélection naturelle de C. Darwin repose sur le fait que les contraintes du milieu favorisent les individus les aptes à se reproduire et donc a transmettre leurs gènes. Par le jeu de ces forces évolutives, de nouvelles espèces apparaissent.
Ces processus génétiques ne sont pas les seuls à l’origine de la diversification des génomes.
En effet, de nombreuses espèces végétales apparaissent suite à l’hybridation de deux espèces parents conduisant à un remaniement chromosomique. Cette hybridation peut être suivie d’une polyploïdisation, ce qui va fortement modifier le contenu génétique des individus en multipliant le nombre des copies des gènes. Ces deux mécanismes peuvent conduire à la naissance de nouvelles espèces.
Les rétrovirus semblent avoir aussi jouer un rôle dans l’évolution des espèces. Ils ont la capacité d’intégrer leur génome dans celui de la cellule hôte. Si cette intégration se fait au niveau des cellules germinales et est stable, les gènes d’origine viraux peuvent être transmis à la descendance et conduire à l’acquisition de nouveaux caractères pouvant favoriser l’apparition d’une nouvelle espèce.
L’organisation et la morphologie des organismes dépend de l’expression spatiale et temporelle des gènes homéotiques. Ce sont des gènes « architectes » responsables de la mise en place des organes et appendices à un endroit donné et à un moment donné du développement embryonnaire.
La mutation de l’un de ces gènes peut conduire à la disparition d’un organe, ou à sa mise en place à un endroit insolite de l’organisme. Elle peut aussi conduire à sa mise en place prématurée ou retardée. C’est l’hétérochronie.
Ainsi, la morphologie et l’organisation de l’organisme seront modifiées au cours du développement embryonnaire, ce qui pourra donner naissance à une nouvelle espèce.
Évalue ce cours !

Des quiz et exercices pour mieux assimiler sa leçon
La plateforme de soutien scolaire en ligne myMaxicours propose des quiz et exercices en accompagnement de chaque fiche de cours. Les exercices permettent de vérifier si la leçon est bien comprise ou s’il reste encore des notions à revoir.

Des exercices variés pour ne pas s’ennuyer
Les exercices se déclinent sous toutes leurs formes sur myMaxicours ! Selon la matière et la classe étudiées, retrouvez des dictées, des mots à relier ou encore des phrases à compléter, mais aussi des textes à trous et bien d’autres formats !
Dans les classes de primaire, l’accent est mis sur des exercices illustrés très ludiques pour motiver les plus jeunes.

Des quiz pour une évaluation en direct
Les quiz et exercices permettent d’avoir un retour immédiat sur la bonne compréhension du cours. Une fois toutes les réponses communiquées, le résultat s’affiche à l’écran et permet à l’élève de se situer immédiatement.
myMaxicours offre des solutions efficaces de révision grâce aux fiches de cours et aux exercices associés. L’élève se rassure pour le prochain examen en testant ses connaissances au préalable.

Des vidéos et des podcasts pour apprendre différemment
Certains élèves ont une mémoire visuelle quand d’autres ont plutôt une mémoire auditive. myMaxicours s’adapte à tous les enfants et adolescents pour leur proposer un apprentissage serein et efficace.
Découvrez de nombreuses vidéos et podcasts en complément des fiches de cours et des exercices pour une année scolaire au top !

Des podcasts pour les révisions
La plateforme de soutien scolaire en ligne myMaxicours propose des podcasts de révision pour toutes les classes à examen : troisième, première et terminale.
Les ados peuvent écouter les différents cours afin de mieux les mémoriser en préparation de leurs examens. Des fiches de cours de différentes matières sont disponibles en podcasts ainsi qu’une préparation au grand oral avec de nombreux conseils pratiques.
Des vidéos de cours pour comprendre en image
Des vidéos de cours illustrent les notions principales à retenir et complètent les fiches de cours. De quoi réviser sa prochaine évaluation ou son prochain examen en toute confiance !





Fiches de cours les plus recherchées


Envie de progresser et de réussir votre année scolaire ?
Testez gratuitement pendant 24h notre plateforme de soutien scolaire !

Un espace dédié aux parents pour suivre les progrès
Tout le programme scolaire du CP à la Terminale
Des profs expérimentés disponibles à la demande par tchat, audio ou vidéo